





Abstract
Objectives
Recent studies have shown that systemic injection of rapamycin can prevent the development of osteoarthritis (OA)-like changes in human chondrocytes and reduce the severity of experimental OA. However, the systemic injection of rapamycin leads to many side effects. The purpose of this study was to determine the effects of intra-articular injection of Torin 1, which as a specific inhibitor of mTOR which can cause induction of autophagy, is similar to rapamycin, on articular cartilage degeneration in a rabbit osteoarthritis model and to investigate the mechanism of Torin 1’s effects on experimental OA.
Methods
Collagenase (type II) was injected twice into both knees of three-month-old rabbits to induce OA, combined with two intra–articular injections of Torin 1 (400 nM). Degeneration of articular cartilage was evaluated by histology using the Mankin scoring system at eight weeks after injection. Chondrocyte degeneration and autophagosomes were observed by transmission electron microscopy. Matrix metallopeptidase-13 (MMP-13) and vascular endothelial growth factor (VEGF) expression were analysed by quantitative RT-PCR (qPCR).Beclin-1 and light chain 3 (LC3) expression were examined by Western blotting.
Results
Intra-articular injection of Torin 1 significantly reduced degeneration of the articular cartilage after induction of OA. Autophagosomes andBeclin-1 and LC3 expression were increased in the chondrocytes from Torin 1-treated rabbits. Torin 1 treatment also reduced MMP-13 and VEGF expression at eight weeks after collagenase injection.
Conclusion
Our results demonstrate that intra-articular injection of Torin 1 reduces degeneration of articular cartilage in collagenase-induced OA, at least partially by autophagy activation, suggesting a novel therapeutic approach for preventing cartilage degeneration and treating OA.
Cite this article: N-T. Cheng, A. Guo, Y-P. Cui. Intra-articular injection of Torin 1 reduces degeneration of articular cartilage in a rabbit osteoarthritis model. Bone Joint Res 2016;5:218–224. DOI: 10.1302/2046-3758.56.BJR-2015-0001.
Article focus
-
To determine the effects of Torin 1 on articular cartilage degeneration in a rabbit osteo-arthritis model by intra-articular injection.
-
Whether intra-articular injection of Torin 1 is accompanied by side effects.
-
Investigate the mechanism of Torin 1’s effects on experimental OA.
Key messages
-
Torin 1 reduced degeneration of articular cartilage and chondrocytes after induction of OA with collagenase injection.
-
The autophagosomes, Beclin-1 and LC3 expression were promoted in chondrocytes from Torin 1-treated rabbits.
-
A reduction in MMP-13 and VEGF expression was also observed in cartilage from Torin 1-treated rabbits after collagenase injection.
Strengths and limitations
-
Intra-articular injection of Torin 1reduces degeneration of articular cartilage in collagenase-induced OA by autophagy activation.
-
The beneficial effect of intra-articular Torin 1 treatment may be due to the inhibition of chondrocyte hypertrophy and angiogenesis.
-
Side effects accompanied by systemic use are not observed.
-
Intra-articular injection of Torin 1 could be a novel therapeutic approach for preventing cartilage degeneration and treating OA.
Introduction
Osteoarthritis (OA) is a common degenerative joint disease causing joint pain, stiffness, deformity and dysfunction in patients. Many factors such as physical, chemical, genetic and ageing factors are thought to be involved in the pathogenesis of OA. Studies have shown that OA is characterised by degeneration of articular cartilage and damage of extracellular matrix (ECM).1 Chondrocytes are the main cells in cartilage and the central feature in cartilage degeneration. Meanwhile, the synthesis and secretion of ECM are regulated by chondrocytes.2 For these reasons, the maintenance of chondrocyte health has been one of the key points for prevention and treatment of OA.
Autophagy, i.e. “self-eating”, is an evolutionarily conserved catabolic process which involves the degradation of damaged organelles and misfolded proteins characterised by the formation of autophagosomes.3 Under stress conditions, autophagy is activated to degrade unnecessary intracellular components for maintaining cell survival.4-6 Autophagy also plays an important role in maintaining metabolic homeostasis,7 and recent studies have revealed that many diseases are associated with dysfunctional autophagy, including ageing,8 neurodegenerative diseases,9 diabetes10 and cardiomyopathy.11 In addition, compromised autophagy isbelievedto be a novel mechanism in the development of OA.12 Based on these findings, autophagy has been suggested to play a protective role in the pathogenesis of OA as it can maintain the health of chondrocytes under conditions of stress.
Mammalian target of rapamycin (mTOR) protein kinase plays a key role in the regulation of cell proliferation, differentiation, motility, metabolism, survival, and autophagy.13 Inhibition of mTOR has been investigated as a potential molecular target for therapeutic intervention of disease.14 Rapamycin is a known inhibitor of mTOR15 and can cause induction of autophagy.16-18 It is a major immunosuppressant used in organ transplantation to induce tolerance in the clinical setting of transplantation.19 Recent studies have revealed that rapamycin can prevent the development of diseases by autophagy enhancement and may represent a novel therapeutic strategy in neurodegenerative diseases,20-22 inflammation and infection,23 spinal cord injury,24,25 renal injury26 and others. In addition, rapamycin has been shown to prevent the expression of OA-like changes in human chondrocytes27 and systemic injection can reduce the severity of experimental OA.28 However, the systemic injection of rapamycin can lead to many side effects, including diarrhoea, weight loss, proteinuria, anaemia, allergy, and increases in serum cholesterol and triglycerides.29 Therefore, intra-articular injection may be an appropriate approach for the clinical use of rapamycin to treat OA.
Similar to rapamycin, Torin 1 is a specific inhibitor of mTOR which can cause induction of autophagy,30 and the effects of Torin 1 on articular cartilage degeneration by intra-articular injection are unknown. The purpose of this study was to determine the effects of intra-articular injection of Torin 1 on articular cartilage degeneration in a rabbit OA model. The results demonstrate that intra-articular injection of Torin 1 reduces degeneration of articular cartilage in collagenase-induced OA by autophagy activation, suggesting a novel therapeutic approach for preventing cartilage degeneration and treating OA.
Materials and Methods
Experimental animals
All studies were performed with the approval of the University Ethics Committee. Male New Zealand, 43-month-old white rabbits were used in this study. The rabbits were anaesthetised with 10% chloral hydrate (2mL / kg body-weight) by intraperitoneal injection and randomly assigned to four groups. In the first group, 0.5 ml saline was injected twice at day one and day four in both knees. In the second group, 0.5 ml saline containing 400 nM Torin 1 was injected twice. In the third group, 0.5 ml saline containing 1.0 mg collagenase (type II) (Sigma-Aldrich, St Louis, Missouri) was injected twice to establish a rabbit OA model as reported.31 In the fourth group, 0.5 ml saline containing 1.0 mg collagenase and 400 nM Torin 1was injected twice. All animals were killed at eight weeks after the initiation of injection and joint tissue was subjected to histological evaluation, transmission electron microscope observation and gene expression analyses.
Histological evaluation and Mankin scores
In all, six knee joints from each group were separated and fixed in 10% neutral buffered formalin, decalcified with 10% EDTA, dehydrated through a series of ethanol washes, and embedded in paraffin. The samples were cut into 5 μm microsections in the sagittal plane and stained with Safranin O. Articular cartilage degeneration was evaluated by two blinded observers using the Mankin scoring system (Table I).32,33
Table I.
Histological evaluation methods of articular cartilage (Mankin scoring system)33
| Articular cartilage |
|---|
| (1) Structure (7 grades) |
| (2) Cell (4 grades) |
| (3) Safranin-O staining (5 grades) |
| (4) Tidemark (2 grades) |
Transmission electron microscopy
We performed transmission electron microscopy to observe chondrocyte degeneration and autophagosomes. Knee joints from each group were fixed with 2.5% glutaraldehyde, decalcified with 10% EDTA, further fixed with 1% OsO4 and dehydrated by a series of ethanol washes. The samples were incubated in isoamyl acetate and embedded in gold palladium. Ultra-thin sections were obtained and observed under a transmission electron microscope (TEM, Hitachi, Japan).
RNA extraction and quantitative RT-PCR (qPCR)
In total, three knee joints from each group were used for qPCR. Total RNA was extracted from cartilage using the TRI zolreagent (Invitrogen,Carlsbad, California)accordingto the manufacturer’s protocol. Quantitative RT-PCRreactions were performed in triplicate using a 7500 Real Time PCR System (ABI, Foster City, California) with SYBR Green qPCR Master Mix (ABI, US). The primers for matrix metallopeptidase-13 (MMP-13), vascular endothelial growth factor (VEGF) and β-actin were designed as follows: MMP-13: forward, 5′-CGACTTCTACCCATTTGA-3′, reverse, 5′-TAGCCTTTGGAACTACTTGTC-3′; VEGF: forward, 5′-CCCACGTCAGAGAGCAACA-3′, reverse, 5′-TCACATCTGCTGTGCTGTAGG-3′; β-actin:forward, 5′-TCGACAACGGCTCCGGCAT-3′,reverse,5′-AAGGTGTGGTGCCAGATTTTC-3′. The amount of target genes, relative to β-actin, was calculated using the 2-ΔΔCT method.
Western blotting
In each group, three knee joints were randomly chosen from the total of nine and then cartilage from these was separated and then pooled as one sample (n = 3). Total protein was extracted and concentrations were determined using a bicinchoninic acid reagent assay (Thermo Fisher Scientific, Waltham, Massachusetts). Total protein was fractionated by Tris-Glycine gels and transferred onto a nitrocellulose membrane. The membrane was blocked with 5% dry milk and incubated with primary antibodies to Beclin-1 (1:1000 dilution), light chain 3 (LC3, Cell Signaling Technology, Danvers, Massachusetts) (1:1000 dilution) and β-actin (Santa Cruz, California) (1:3000 dilution). The blots were then developed using a horseradish peroxidase-conjugated secondary antibody (Beijing Zhongshan Golden Bridge Biotechnology, Beijing, China) and enhanced chemiluminescence substrate (Thermo). The WB data were quantified by GEL – DOC 2000 UV GEL imaging system (Bio-Rad, Hercules, California).
Statistical analysis
The data were expressed as the mean and standard deviation (sd). Statistical analysis was performed using SPSS statistical software 19.0 (SPSS Inc., Chicago, Illinois). The Student’s t-test was used for direct comparisons between the two groups. One-way analysis of variance was used for multiple comparisons between independent groups. A p-value less than 0.05 was considered statistically significant.
Results
Intra-articular injection of Torin 1 reduced degeneration of articular cartilage in collagenase-induced OA
After intra-articular injection, side effects such as diarrhoea and allergy (monitored by observation) weight loss, proteinuria (checked by urine test), anaemia and increases in serum cholesterol and triglycerides (checked by blood test) and wound infection were not observed in any of the experimental groups. There were no degenerative changes in the cartilage in the knees from control rabbits that were treated with saline or Torin 1;the surface of the cartilage was smooth and regular. Histological evaluation showed that the saline-treated rabbits had loss of cartilage surface and Safranin O staining, and the presence of lesions extending into the radial zone at eight weeks after collagenase injection. In contrast, in Torin 1-treated rabbits, the loss of Safranin O staining and the loss of chondrocytes was significantly less in the controls in the transitional zone at eight weeks after collagenase injection, and the cartilage surface was preserved. In collagenase-induced rabbits, histological evaluation demonstrated that there was significantly less degeneration of articular cartilage in the rabbits treated with intra-articular injection of Torin 1 by at eight weeks compared with the rabbits treated with saline (Fig. 1a). The extent of OA was evaluated by the Mankin scoring system and presented as the average of summed scores (the higher the score, the greater the extent of OA) (Table II, Fig. 1b). The scores obtained showed that saline-treated rabbits had a significantly higher Makin score than Torin 1-treated rabbits at eight weeks after induction of OA with collagenase injection (p < 0.001). These results demonstrated that intra-articular injection of Torin 1 reduced degeneration of articular cartilage in collagenase-induced OA.
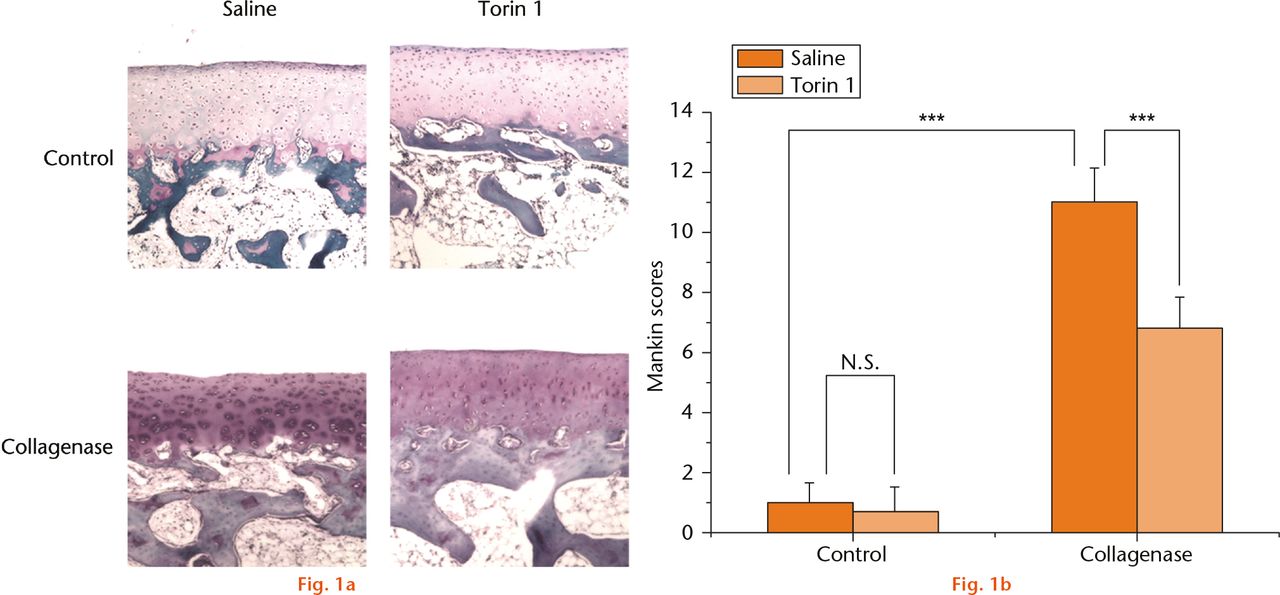
Fig.
Histological evaluation of osteoarthritis: (a) there were no degenerative changes of the cartilage in the knees from control rabbits that were treated with saline or Torin 1. The saline-treated rabbits had loss of cartilage surface, Safranin O staining, and had lesions extending into the radial zone at eight weeks after collagenase injection. In contrast, Torin 1-treated rabbits had abrogated loss of Safranin O staining and chondrocyte disappearance in the transitional zone at eight weeks after collagenase injection, and the cartilage surface was preserved. b) The Mankin scores showed that saline-treated rabbits had a significantly higher score than Torin 1-treated rabbits at eight weeks after induction of OA with collagenase injection. In saline-treated rabbits, the score of the collagenase-induced group was significantly higher than the control group. ***p < 0.001 (Magnification, ×100).
Table II.
The mean Mankin scores
| Mankin scores | Mean (sd) (n = 6) |
|---|---|
| Saline | 1.00 (0.63) |
| Saline + Torin 1 | 0.67 (0.82) |
| Collagenase | 10.83 (1.17) |
| Collagenase + Torin 1 | 6.67 (1.03) |
Intra-articular injection of Torin 1 increased autophagosomes and reduced degeneration of chondrocytes in collagenase-induced OA
The autophagosomes and degeneration of chondrocytes were observed by TEM (Fig. 2). Many more autophagic vacuoles were observed in chondrocytes from Torin 1-treated rabbits than from saline-treated rabbits; the autophagy was promoted by treatment of Torin 1. In control rabbits treated with saline or Torin 1, chondrocytes located in the lacunae contained autophagic vesicles and were not degenerated. In addition, Golgi apparatus and abundant rough endoplasmic reticulum (RER) were also observed. TEM observation showed that chondrocytes from saline-treated rabbits were condensed with absent nuclei and several autophagosomes in the cytoplasm at eight weeks after collagenase injection, and some cell debris were seen in the lacunae. In contrast, chondrocytes from Torin 1-treated rabbits had nuclei and condensed chromatin at eight weeks after collagenase injection. TEM observation demonstrated less degeneration of chondrocytes in rabbits treated with Torin 1 compared with rabbits treated with saline at eight weeks after induction of OA with collagenase injection. These results demonstrated that intra-articular injection of Torin 1 increased autophagosomes and reduced degeneration of chondrocytes in collagenase-induced OA.
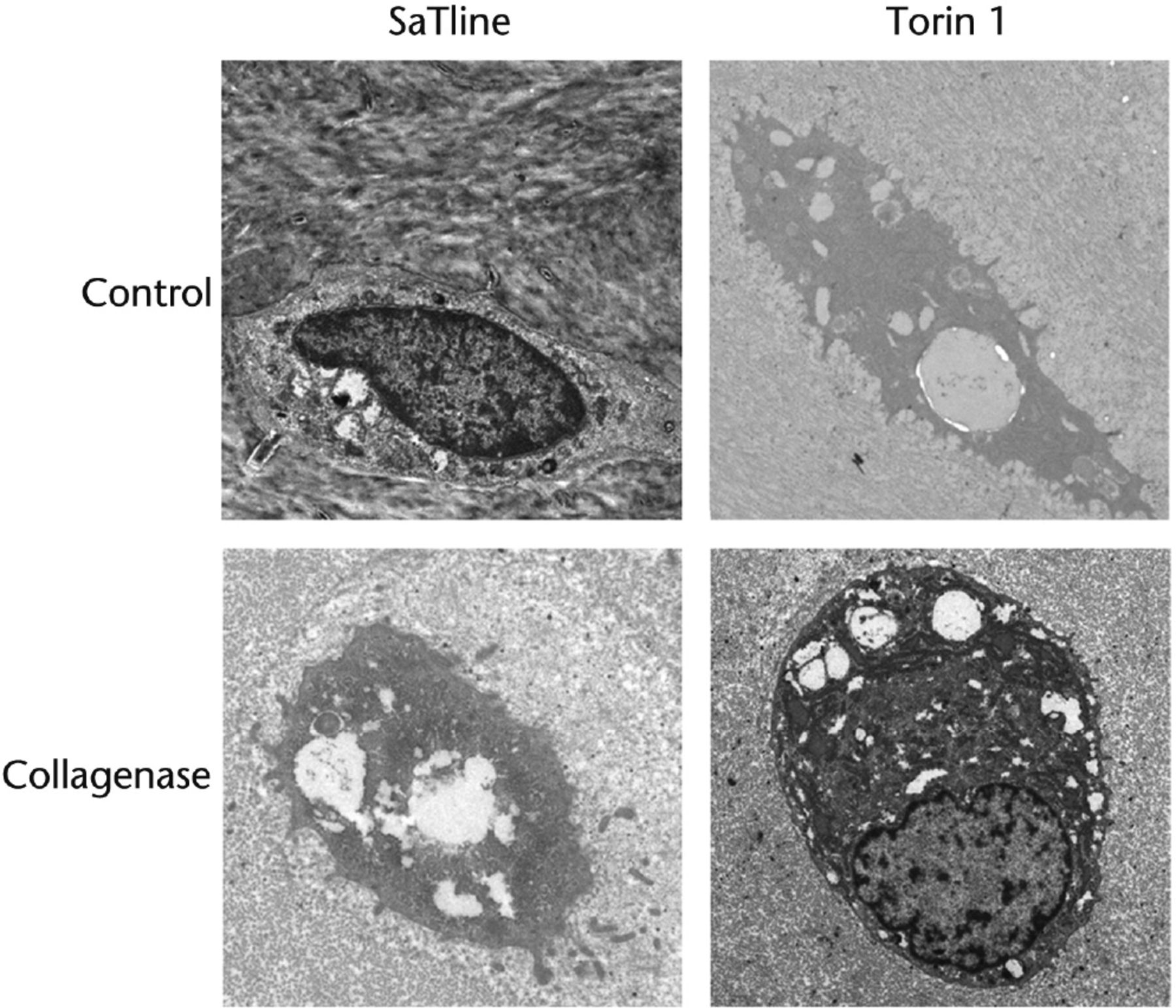
Fig. 2
TEM observation. Many more autophagic vacuoles were observed in chondrocytes from Torin 1-treated rabbits than from saline-treated rabbits. In control rabbits treated with saline or Torin 1, chondrocytes located in the lacunae contained autophagic vesicles and were not degenerated. In addition, Golgi apparatus and abundant rough endoplasmic reticulum (RER) were also observed. Chondrocytes from saline-treated rabbits were condensed with absent nuclei and several autophagosomes in the cytoplasm at eight weeks after collagenase injection, and some cell debris were seen in the lacunae. In contrast, chondrocytes from Torin 1-treated rabbits had nuclei and condensed chromatin at eight weeks after collagenase injection (Bar, 1μm).
Intra-articular injection of Torin 1 decreased MMP-13 and VEGF expression in collagenase-induced OA
Reports have shown that the expression of MMP-13 and VEGF is involved in the pathological process of OA.34,35 To study the mechanism whereby Torin 1 can prevent experimental OA, the expression of MMP-13 and VEGF was evaluated by qPCR (Fig. 3). The expression of MMP-13 and VEGF showed a significant increase in the saline-treated rabbits at eight weeks after collagenase injection compared with control knees (p < 0.001). Torin1 treatment decreased the expression of MMP-13 and VEGF when compared with the saline-treated rabbits at eight-weeks post collagenase injection (p < 0.001) (Figs 3a and 3b), suggesting that intra-articular injection of Torin 1 may reduce degeneration of articular cartilage and chondrocytes through a decrease in MMP-13 and VEGF expression.
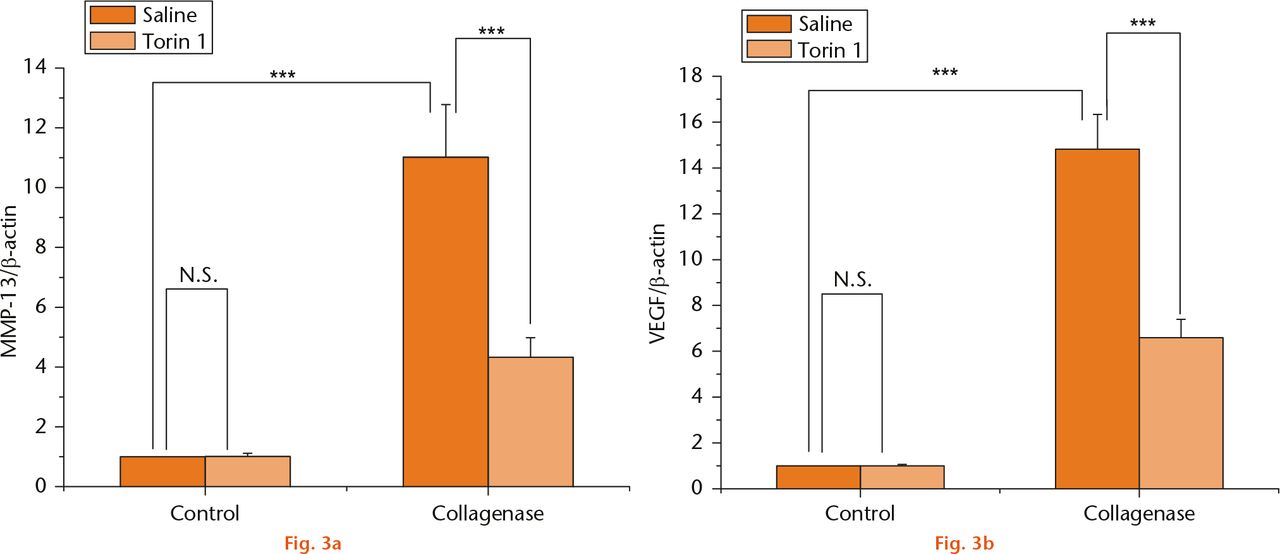
Fig.
MMP-13 and VEGF expression (a) and (b): the mRNA expression of MMP-13 and VEGF revealed a significant increase in the saline-treated rabbits at eight weeks after collagenase injection compared with control knees. Torin 1 treatment decreased the expression of MMP-13 and VEGF when compared with saline-treated rabbits at eight-week time points post collagenase injection ***p < 0.001.
Intra-articular injection of Torin 1 increased Beclin-1 and LC3 expression in collagenase-induced OA
To determine whether intra-articular injection of Torin 1 modulates autophagy, the effects of Torin 1 on Beclin-1 and LC3 expression were evaluated by Western blotting (Fig. 4a). The expression ofBeclin-1 and LC3 was significantly decreased in saline-treated rabbits at eight weeks post-collagenase injection compared with saline-treated rabbits of the control group (p < 0.001). Torin 1 treatment increased Beclin-1 and LC3 expression in control rabbits compared with those treated with saline at eight weeks (p = 0.002 and p = 0.001). Similarly, the expression of Beclin-1 and LC3 was increased in Torin 1-treated rabbits compared with those treated with saline at eight weeks after collagenase injection (p < 0.001) (Figs 4b and 4c). These results further demonstrated that intra-articular injection of Torin 1 promoted chondrocyte autophagy.
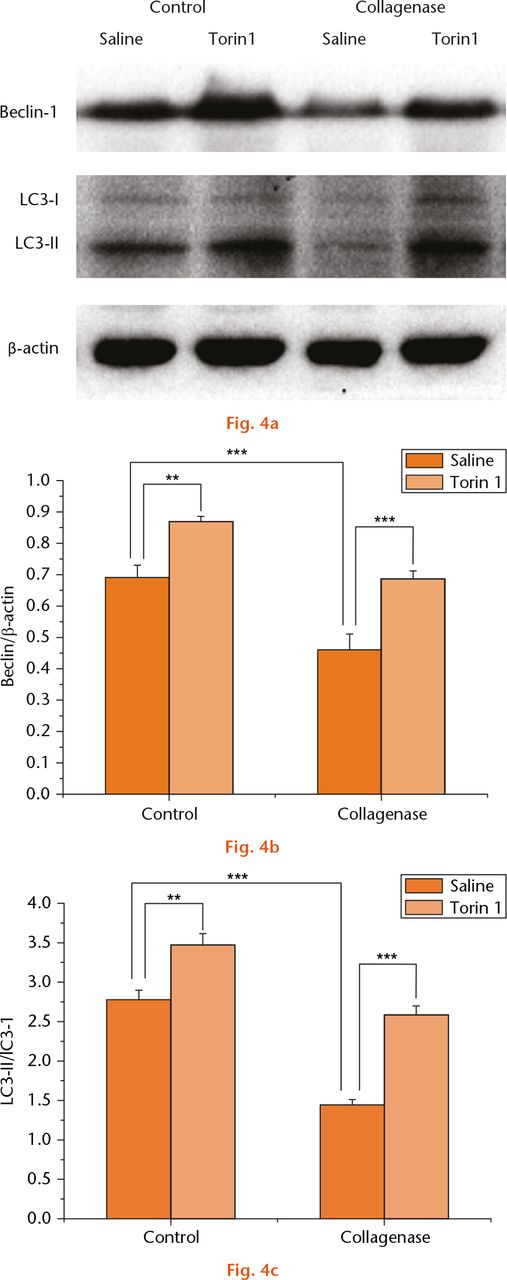
Fig.
Beclin-1 and LC3 expression: a) the expression of Beclin-1 and LC3 evaluated by Western blotting (WB), (b) and (c). The expression of Beclin-1 and LC3 was significantly decreased in saline-treated rabbits at eight weeks post-collagenase injection compared with saline-treated rabbits of the control group. Torin 1 treatment increased Beclin-1 and LC3 expression in control rabbits compared with those treated with saline at eight weeks. Similarly, the expression of Beclin-1 and LC3 was increased in Torin 1-treated rabbits compared with those treated with saline at eight weeks after collagenase injection. **p < 0.01, ***p < 0.001. The WB data were quantified by GEL – DOC 2000 UV GEL imaging system.
Discussion
Effective therapies to prevent the pathological progression of OA continue to be critical topics in clinical medicine. Recently, rapamycin has been used to treat OA by systemic injection and the results show that it can reduce the severity of experimental OA in a mouse model.28 However, systemic injection of rapamycin is accompanied by many side effects including diarrhoea, weight loss, proteinuria, anaemia, allergy, and increases in serum cholesterol and triglycerides.29 None of these side effects were observed in this study by intra-articular injection of Torin 1, suggesting that intra-articular injection may be an appropriate approach for the clinical use of Torin 1 to treat OA.
Previously, a rabbit model of OA was established by intra-articular injection of collagenase.31 The results demonstrated degeneration of articular cartilage after collagenase injection. The current study has shown that the intra-articular injection of Torin 1 significantly reduces the degeneration of articular cartilage in this collagenase model.
To study the mechanism of Torin 1’s effect on experimental OA, we focused on the expression of MMP-13 and VEGF. MMP-13 is involved in the progression of OA and is a widely used hypertrophic chondrocyte marker.36 Studies have revealed that hypertrophic chondrocytes promote the development of OA, and inhibition chondrocyte hypertrophy may be a potential therapeutic target for OA.37 Meanwhile, mTOR is related to chondrocyte hypertrophy38 and rapamycin can reduce hypertrophic changes of chondrocytes through mTOR inhibition.39 In addition, angiogenesis has been suggested to play an important role in the degeneration of articular cartilage after OA, and the level of VEGF in OA cartilage is increased.34 Inhibition of VEGF has been shown to decrease angiogenesis and prevent OA-like changes in experimental OA.40 mTOR is also related to angiogenesis and the use of rapamycin leads to a decrease in VEGF expression.41 In this study, we found that Torin 1 treatment decreased the expression of MMP-13 and VEGF when compared with rabbits treated with saline after collagenase injection, suggesting that intra-articular injection of Torin 1 reduced degeneration of articular cartilage and chondrocytes through the inhibition of chondrocyte hypertrophy and angiogenesis.
mTOR interfaces with many aspects of cellular processes, such as transcription, translation, protein turnover and cytoskeletal organisation.42 Phosphatidylinositol 3 kinase (PI3K) and protein kinase B (PKB or Akt) have been implicated in the activation of mTOR. The PI3K/Akt/mTOR signaling pathway regulates cell growth and survival in response to nutrients and stresses.43 Studies have revealed that the PI3K/Akt/mTOR signaling pathway also regulates the modulation of autophagy,44,45 a degradation process that delivers damaged organelles and misfolded proteins to lysosomes, and then the contents are degraded by lysosome enzymes for recycling. A recent study has demonstrated that the downstream modulation of autophagy by mTOR in articular cartilage homeostasis protects mice from OA.46 Autophagy modulates OA-related gene expression in human chondrocytes, and increased autophagy is an adaptive response to protect cells from stresses,27 while autophagy is significantly reduced in late-stage OA.12 In the current study, we found that autophagosomes and the expression of Beclin-1 and of LC3 (autophagy marker) were significantly decreased in degenerating cartilage at eight weeks after collagenase injection compared with normal cartilage, which further demonstrates that autophagy loss partially results in the degeneration of articular cartilage after induction of OA with collagenase injection. Moreover, our results also determined that intra-articular injection of Torin 1 reduced degeneration of articular cartilage, while autophagosomes and the expression of Beclin-1 and of LC3 were increased.
In conclusion, our results demonstrated that Torin 1 reduced degeneration of articular cartilage after induction of OA with collagenase injection. Autophagosomes and Beclin-1 and LC3 expression were upregulated in chondrocytes from Torin 1-treated rabbits. A reduction in MMP-13 and VEGF expression was also observed in cartilage from Torin 1-treated rabbits after collagenase injection. These observations indicate that intra-articular injection of Torin 1reduces degeneration of articular cartilage in collagenase-induced OA by autophagy activation, while side effects accompanied by systemic use are not observed, which suggests that intra-articular injection of Torin 1 could be a novel therapeutic approach for preventing cartilage degeneration and treating OA.
Funding Statement
None declared.
ICMJE conflict of interest
None declared.
References
1 Lahm A , KaschR, MrosekE, et al.. Semiquantitative analysis of ECM molecules in the different cartilage layers in early and advanced osteoarthritis of the knee joint. Histol Histopathol2012;27:609-615.CrossrefPubMed Google Scholar
2 Kronenberg HM . Developmental regulation of the growth plate. Nature2003;423:332-336.CrossrefPubMed Google Scholar
3 Ryter SW , CloonanSM, ChoiAM. Autophagy: a critical regulator of cellular metabolism and homeostasis. Mol Cells2013;36:7-16.CrossrefPubMed Google Scholar
4 Urbanek T , KuczmikW, Basta-KaimA, GabryelB. Rapamycin induces of protective autophagy in vascular endothelial cells exposed to oxygen-glucose deprivation. Brain Res2014;1553:1-11.CrossrefPubMed Google Scholar
5 Kroemer G , MariñoG, LevineB. Autophagy and the integrated stress response. Mol Cell2010;40:280-293.CrossrefPubMed Google Scholar
6 Lee JS , LeeGM. Rapamycin treatment inhibits CHO cell death in a serum-free suspension culture by autophagy induction. Biotechnol Bioeng2012;109:3093-3102.CrossrefPubMed Google Scholar
7 Jin Y , TanakaA, ChoiAM, RyterSW. Autophagic proteins: new facets of the oxygen paradox. Autophagy2012;8:426-428.CrossrefPubMed Google Scholar
8 Markaki M , TavernarakisN. Metabolic control by target of rapamycin and autophagy during ageing - a mini-review. Gerontology2013;59:340-348.CrossrefPubMed Google Scholar
9 Carloni S , BuonocoreG, LonginiM, ProiettiF, BalduiniW. Inhibition of rapamycin-induced autophagy causes necrotic cell death associated with Bax/Bad mitochondrial translocation. Neuroscience2012;203:160-169.CrossrefPubMed Google Scholar
10 Cuervo AM . Autophagy and aging: keeping that old broom working. Trends Genet2008;24:604-612.CrossrefPubMed Google Scholar
11 Nair S , RenJ. Autophagy and cardiovascular aging: lesson learned from rapamycin. Cell Cycle2012;11:2092-2099.CrossrefPubMed Google Scholar
12 Caramés B , TaniguchiN, OtsukiS, BlancoFJ, LotzM. Autophagy is a protective mechanism in normal cartilage, and its aging-related loss is linked with cell death and osteoarthritis. Arthritis Rheum2010;62:791-801.CrossrefPubMed Google Scholar
13 Li H , JinX, ZhangZ, XingY, KongX. Inhibition of autophagy enhances apoptosis induced by the PI3K/AKT/mTor inhibitor NVP-BEZ235 in renal cell carcinoma cells. Cell BiochemFunct2013;31:427-433.CrossrefPubMed Google Scholar
14 Cao C , SubhawongT, AlbertJM, et al.. Inhibition of mammalian target of rapamycin or apoptotic pathway induces autophagy and radiosensitizes PTEN null prostate cancer cells. Cancer Res2006;66:10040-10047. Google Scholar
15 Wu L , FengZ, CuiS, et al.. Rapamycin upregulates autophagy by inhibiting the mTOR-ULK1 pathway, resulting in reduced podocyte injury. PLoS One2013;8:e63799.CrossrefPubMed Google Scholar
16 Kalamidas SA , KondomerkosDJ, KotoulasOB, HannAC. Electron microscopic and biochemical study of the effects of rapamycin on glycogen autophagy in the newborn rat liver. Microsc Res Tech2004;63:215-219.CrossrefPubMed Google Scholar
17 Tanemura M , SagaA, KawamotoK, et al.. Rapamycin induces autophagy in islets: relevance in islet transplantation. Transplant Proc2009;41:334-338.CrossrefPubMed Google Scholar
18 Li X , WuD, ShenJ, ZhouM, LuY. Rapamycin induces autophagy in the melanoma cell line M14 via regulation of the expression levels of Bcl-2 and Bax. Oncol Lett2013;5:167-172.CrossrefPubMed Google Scholar
19 Xiong A , DuanL, ChenJ, et al.. Flt3L combined with rapamycin promotes cardiac allograft tolerance by inducing regulatory dendritic cells and allograft autophagy in mice. PLoS One2012;7:e46230.CrossrefPubMed Google Scholar
20 Majumder S , RichardsonA, StrongR, OddoS. Inducing autophagy by rapamycin before, but not after, the formation of plaques and tangles ameliorates cognitive deficits. PLoS One2011;6:e25416. Google Scholar
21 Cai Z , ZhaoB, LiK, et al.. Mammalian target of rapamycin: a valid therapeutic target through the autophagy pathway for Alzheimer’s disease?J Neurosci Res2012;90:1105-1118.CrossrefPubMed Google Scholar
22 Floto RA , SarkarS, PerlsteinEO, et al.. Small molecule enhancers of rapamycin-induced TOR inhibition promote autophagy, reduce toxicity in Huntington’s disease models and enhance killing of mycobacteria by macrophages. Autophagy2007;3:620-622. Google Scholar
23 Abdulrahman BA , KhweekAA, AkhterA, et al.. Autophagy stimulation by rapamycin suppresses lung inflammation and infection by Burkholderiacenocepacia in a model of cystic fibrosis. Autophagy2011;7:1359-1370. Google Scholar
24 Sekiguchi A , KannoH, OzawaH, YamayaS, ItoiE. Rapamycin promotes autophagy and reduces neural tissue damage and locomotor impairment after spinal cord injury in mice. J Neurotrauma2012;29:946-956.CrossrefPubMed Google Scholar
25 Chen HC , FongTH, HsuPW, ChiuWT. Multifaceted effects of rapamycin on functional recovery after spinal cord injury in rats through autophagy promotion, anti-inflammation, and neuroprotection. J Surg Res2013;179:e203-e210.CrossrefPubMed Google Scholar
26 Xiao T , GuanX, NieL, et al.. Rapamycin promotes podocyte autophagy and ameliorates renal injury in diabetic mice. Mol Cell Biochem2014;394:145-154.CrossrefPubMed Google Scholar
27 Sasaki H , TakayamaK, MatsushitaT, et al.. Autophagy modulates osteoarthritis-related gene expression in human chondrocytes. Arthritis Rheum2012;64:1920-1928.CrossrefPubMed Google Scholar
28 Caramés B , HasegawaA, TaniguchiN, et al.. Autophagy activation by rapamycin reduces severity of experimental osteoarthritis. Ann Rheum Dis2012;71:575-581.CrossrefPubMed Google Scholar
29 Cravedi P , RuggenentiP, RemuzziG. Sirolimus to replace calcineurin inhibitors? Too early yet.Lancet2009;373:1235-1236.CrossrefPubMed Google Scholar
30 Francipane MG , LagasseE. Selective targeting of human colon cancer stem-like cells by the mTOR inhibitor Torin-1. Oncotarget2013;4:1948-1962.CrossrefPubMed Google Scholar
31 Kikuchi T , SakutaT, YamaguchiT. Intra-articular injection of collagenase induces experimental osteoarthritis in mature rabbits. Osteoarthritis Cartilage1998;6:177-186.CrossrefPubMed Google Scholar
32 Mankin HJ , JohnsonME, LippielloL. Biochemical and metabolic abnormalities in articular cartilage from osteoarthritic human hips. III. Distribution and metabolism of amino sugar-containing macromolecules. J Bone Joint Surg [Am]1981;63-A:131-139.PubMed Google Scholar
33 Mankin HJ , DorfmanH, LippielloL, et al.. Biochemical and metabolic abnormalities in articular cartilage from osteoarthritic human hips. II. Correlation of morphology with biochemical and metabolic data. J Bone Joint Surg [Am]1971;53-A:523-531. Google Scholar
34 Wei T , KulkarniNH, ZengQQ, et al.. Analysis of early changes in the articular cartilage transcriptisome in the rat meniscal tear model of osteoarthritis: pathway comparisons with the rat anterior cruciate transection model and with human osteoarthritic cartilage. Osteoarthritis Cartilage2010;18:992-1000.CrossrefPubMed Google Scholar
35 Yang L , GuoA, GuJ C. c-Jun N-terminal kinase and nuclear factor κB mediate nitric oxide-induced expression of matrix metalloproteinase-13. Int Orthop2011;35:1261-1266.CrossrefPubMed Google Scholar
36 Alvarez J , BalbinM, SantosF, et al.. Different bone growth rates are associated with changes in the expression pattern of types II and X collagens and collagenase 3 in proximal growth plates of the rat tibia. J Bone Miner Res2000;15:82-94.CrossrefPubMed Google Scholar
37 van der Kraan PM , van den BergWBM. Chondrocyte hypertrophy and osteoarthritis: role in initiation and progression of cartilage degeneration?Osteoarthritis Cartilage2012;20:223-232.CrossrefPubMed Google Scholar
38 Park IH , ErbayE, NuzziP, ChenJ. Skeletal myocyte hypertrophy requires mTOR kinase activity and S6K1. Exp Cell Res2005;309:211-219.CrossrefPubMed Google Scholar
39 Alvarez-García O , García-LópezE, LoredoV, et al.. Rapamycin induces growth retardation by disrupting angiogenesis in the growth plate. Kidney Int2010;78:561-568.CrossrefPubMed Google Scholar
40 Matsumoto T , CooperGM, GharaibehB, et al.. Cartilage repair in a rat model of osteoarthritis through intraarticular transplantation of muscle-derived stem cells expressing bone morphogenetic protein 4 and soluble Flt-1. Arthritis Rheum2009;60:1390-1405.CrossrefPubMed Google Scholar
41 Guba M , von BreitenbuchP, SteinbauerM, et al.. Rapamycin inhibits primary and metastatic tumor growth by antiangiogenesis: involvement of vascular endothelial growth factor. Nat Med2002;8:128-135.CrossrefPubMed Google Scholar
42 Sabatini DM . mTOR and cancer: insights into a complex relationship. Nat Rev Cancer2006;6:729-734.CrossrefPubMed Google Scholar
43 Cantley LC . The phosphoinositide 3-kinase pathway. Science2002;296:1655-1657. Google Scholar
44 Chang YY , JuhászG, Goraksha-HicksP, et al.. Nutrient-dependent regulation of autophagy through the target of rapamycin pathway. Biochem Soc Trans2009;37:232-236.CrossrefPubMed Google Scholar
45 Kondomerkos DJ , KalamidasSA, KotoulasOB, HannAC. Glycogen autophagy in the liver and heart of newborn rats. The effects of glucagon, adrenalin or rapamycin.Histol Histopathol2005;20:689-696. Google Scholar
46 Zhang Y , VasheghaniF, LiYH, et al.. Cartilage-specific deletion of mTOR upregulates autophagy and protects mice from osteoarthritis. Ann Rheum Dis2015;74:1432-1440.CrossrefPubMed Google Scholar
